




肽类神经递质的共存与受体

厦门心理咨询-德仁心心理咨询机构:肽类神经递质的共存与受体
编辑:徐蒙蒙
肽类神经递质
已知许多肽记内分泌细胞分泌的激素、包括一些脑内神经内分泌区域的神经元所分泌的激素。近来,发现许多肽类物质起着神经递质的作用,且常常与小分子神经质一同释放。
神经肽的生物学活性取决于其氨基酸序列。也就是说,神经肽的前体的分子常常比活性产物大许多倍,并能产生不止一种的神经肽。这些肽类产物可以单独存在于不同的突触囊泡中,故神经肽对突触后神经元的作用是很复杂的。肽类递质已被证明同人的情绪调节、疼痛感受和紧张反应密切相关。
P物质是最早发现的神经肽物质,含有11个氨基酸。在人海的脑马和新皮质内含量很高;P物质也有周围神经中口径较小的C纤维所释放,与传导疼痛和温度感觉功能有关。作为感觉性神经递质,P物质的的传递在脊椎中可被中间神经释放的阿片肽所抑制。
阿片肽类是在脑内发现的有类似吗啡作用的肽类物质,而吗啡是有镇痛剂。其作用机制是与存在于中枢及周围神经系统许多部位、传导疼痛信息通路中的特殊受体有关。现已发有超过20种的阿片肽类物质,可分成三类:内啡肽、脑啡肽和强啡狀。每种狀都来自一种没有活性的前体肽、分属三种不同的基因,这些前体的加工有明显的组织特异性。阿片肽类递质在脑内分布广泛,总的起镇静作用,故可在脑内注射时起镇痛作用,是针刺能引起麻醉的机制。但是,重复使用内啡狀可引起成瘾或耐受性。
此外,经常提到的神经肽类还有神经垂体源性肽类(如加压素、催产素)血管活性肠肽、胰岛素类肽生长抑素、胃泌素和但那收缩素等等。现在已发现的神经肽欧50种以上,对不同神经元起着兴奋或抑制作用。
关于递质共存
近来发现神经系统许多部位的单一神经元可以释放不止一种的神经递质,它们在神经元中呈现递质共存现象。这一般指传统小分子递质与神经肽的共存,目前也发现有两种小分子递质如5-羟色胺与GABA存在于同一个神经元中的情况。两种或更多的递质共存也呈现多种搭配关系,如脑啡肽既可以在交感神经系统中的某些去甲基肾上腺素能神经元中存在,也可以在脑内某些5 -羟色胺能神经元中呈现。这就说明脑内神经元突触传递具有髙度复杂性。
递质共存的功能意义目前还不很清楚,有研究证明神经肽是在神经元出琬明显功能活动期间释放,而传统的小分子递质的释放则与神经冲动传导有关。例如,研究发现支配唾液腺的副交感神经元释放乙酰胆碱和血管活性肠肽。其中,乙酰胆碱作用在唾液腺分泌细胞的毒蕈碱受体从而刺激细胞分泌黏液;共同释放的血管活性肠肽则作用在小动脉的受体,造成血流增加和体液分泌。同样,在脑内发现投射到皮质边缘系统的多巴胺能神经元与胆囊收缩素之间有共存,但投射到纹状体的多巴胺能神经元则没有这样的共存现象,说明胆囊收缩素只调节皮质边缘系统中的多巴胺释放而对纹状体的多巴胺不起调节作用。这种选择性的递质共存现象可被用来考虑,使用与胆囊收缩素受体有关的药物来治疗涉及皮质边缘系统中的多巴胺能神经元的精神障碍疾患。
什么是受体?
在神经科学中有关脑内信息加工和包括神经精神在内的神经活性物质作用部位的大量进展都取决于对脑内受体的深入研究。神经递质之所以能唤起突触后细胞的电反应是由于其能和突触后细胞膜上的各种蛋白相结合的缘故。这些蛋白质称为递质受体,它们与递质结合后,可以通过打开或关闭突触后膜上的离子通道而引出电信号。某种递质在突触后神经元上能引起兴奋或抑制效应完全取决于这种递质所作用的离子通道的种类不同。受体一般分两大类;一类受体本身就是离子通道的一种;另一类受体则是与离子通道相分离的另一组分子。前一类受体称为离子趋向型受体, —般引起快速突触后反应,持续仅数毫秒;后一类受体称代谢趋向型受体,它们引起较慢的突触后效应。代谢趋向型受体对突触后离子通道是间接地通过一种传感器分子——G蛋白分子实现的,G蛋白的活动可以直接改变离子通道的特性,也可以间接地通过激活细胞内的第二信使途径再调节通道的活动。
神经递质与受体的结合具有特异性、亲合性、可逆性和饱和性(由于受体数量是有限的)的特点。根据这些特点,人们研究受体可通过使用某种放射性配体来 对受体进行特异性标记。如果这种放射性配体与受体之间产生出足够高程度的亲合力,那么,放射性配体就会与受体发生较长时间的结合,以致可将这种已经带有放射性的结合体分离出来。用这种办法,就可以研究受体与递质相互作用的特性及其在神经系统中的定位。例如,如果发现某种药物或制剂可以阻碍放射性配体与受体的特异结合,则说明这种药物或制剂可能属于与该受体有关的递质相似物质。同时,这种配体与受体相结合的方法也可以用作确定受体识别部位的三维分子构型。这些办法还可以与活体生理或行为学实验结合起来研究,即可以先将某种与递质相关的药物或制剂用于活体或行为观察,再取动物组织匀浆观察受体与之相互作用的状况,从而获得相关性结果。例如,曾有人用此方法看到抗精神病药与多巴胺受体结合程度与其治疗精神分裂症效果之间存在有一定相关现象。
由于放射性配体与受体之间的结合往往相当紧密,因此,人们可以用放射自显影技术来观察受体在脑内分布的状况。也就是说,可用脑不同部位的薄切片与含有放射性配体的缓冲液一起孵育一定时间后,再用缓冲液冲洗掉游离的和与组织结合不紧密的放射性配体,这样就保留了与脑组织内受体紧密结合的放射性配体,通过乳胶显影照相,析出的银颗粒分布范围即代表了受体的精确部位。同样,使用当前更新一代的仪器设备,如正电子放射显影技术,在电子计算机的控制下观察三维空间,可以准确看到包括人类在内的活体动物脑内不同递质受体的分布情况。例如,使用一种用放射性碳(11C)标记的能发射正电子的化合物,由于它能高亲和性地与多巴胺受体D2相结合,就可以通过正电子扫描成像技术看到人脑内D2受体的分布情况。
前面已经提到,受体可以幵放或关闭突触后离子通道,这些功能由两类受体蛋白通过两种方式来实现。其中,离子趋向型受体分子含有两个机能基团:一个位于细胞外,专门与受体相结合;另一个骑跨在膜上,本身构成离子通道。由于这两个基团属于同一分子,因此,离子趋向型受体又称配体门控离子通道。另— 类代谢趋向型受体分子本身不包含离子通道,它们与通道之间存在有G蛋白,因此,也叫做G蛋白偶联受体,这类受体属于单体蛋白质,—部分位于细胞外与递质结合,一部分在细胞内与G蛋白结合。G蛋白可以直接作用到离子通道上,也可以先作用在其他效应蛋白(如某种酶)上,再间接地对禽子通道产生影响。因此,G蛋白起着传感器的作用。分别属于上述两类受体中的各种受体所唤起的突触后电位持续时间大不相同,可以从几毫秒到数天之久。配体门控离子通道引起的反应均较快,如乙酰胆碱能的神经肌肉接点引起的兴奋性突触后电位只在1毫秒之内;而代谢趋向型受体中,G蛋白直接作用在通道上的为几百毫秒,而G蛋白间接作用在通道上的可持续数秒、数小时甚至更长。需要指出的是,某种递质可以同时引起两类受体的活动,即同时在一个突触中产生快和慢的突触后电位。但无论何种受体,在递质持续存在的情况下都可产生去敏感现象,这就可以影响突触后电位所持续的时间。
研究最多的配体门控离子通道是烟碱乙酰胆碱受体,这种受体先发现于神经肌肉接点,其命名是由于烟碱也能与之结合的缘故。许多生物毒性物质可特异性地阻断这一受体,如一种蛇毒就可以用来分离和纯化。这样,就知道构成这种受体的蛋白质是由5个亚单位围绕一个中央孔洞所组成。每个亚单位都含有四个跨膜单位,组成离子通道部分,同时各自还有一个长的细胞外区域即同乙酰胆碱结合的基团。针对谷氨酸递质的受体中,NMDA (N-甲基-D-天门冬氨酸)受体和AMPA (a-氨基-3_经基-5-甲基-4-isoxazole_丙酸)/海人酸受体属于配体门控离子通道。AMPA/海人酸受体在谷氨酸递质作用下引起最快速的兴奋性突触后电位。这种受体也由若干蛋白质亚单位组成,这些亚单位所形成更多种类型的受体,而由亚单位所形成的中心孔洞可以允许Na+和K+离子通过。同样,NMDA受体也是由多个选择性离子通道亚单位所构成,但其具有特别的性能,即这些通道除能允许单价离子(如Na+和K+)通过外还可以允许Ca2+离子进入。因此,NMDA受体所产生的兴奋性突触后电位可以增加突触后神经元内的钙离子浓度,也就是说,钙离子浓度变化就在细胞内起到了第二信使的作用。另外,NMDA受体仅在突触后细胞去极化而消除细胞外镁离子(Mg2+)的阻断时才能允许离子通过,这一机制涉及到成年大脑对信息的储存功能,目前对此巳有不少研究。
以GABA为递质的受体有两类:其中一类属配体门控离子通道,即GABAa受体。由于此类通道可以通过带负电的氯离子,故其具有抑制功能。这类受体是临床上多种药物的作用部位。例如,安定和甲氨二氮罩这种精神安定药物可以与GABAa受体的a和b亚单位相结合;而巴比妥一类的催眠药则与其r亚单位相结合。酒精也能作用在GABA受体上,可能因干扰了抑制性突触传递而产生某种中毒后表现的行为。
某些5-羟色胺受体(如5-HT3受体)和嘌呤能受体(如P2x受体)也属于配体门控离子通道。它们都是非选择性离子通道,常能允许Ca2+进入。
就代谢趋向型受体而论,目前巳鉴定出超过100种不同的受体。这包括多种小分子递质受体,如谷胺酸的mGluRl〜6;肾上腺素和去甲基肾上腺素的a1、a2、b1和b2;匕和乙酰胆碱的ml〜m5;多巴胺的D1^D5;GABA的GABAb以及所有神经肽的受体。此类受体仅由一个蛋白分子构成,但包含有7个跨膜单位,其中至少有4个在细胞外与递质相结合,两个则与G蛋白结合。受到受体作用的G蛋白有三部分组成(a、b和r),因此,也称为三供体G蛋白,其中a亚单位可以与鸟嘌呤呤核苷酸(如GTP和GDP)相结合。当a亚单位与GDP相结合时,a亚单位还能与b和r
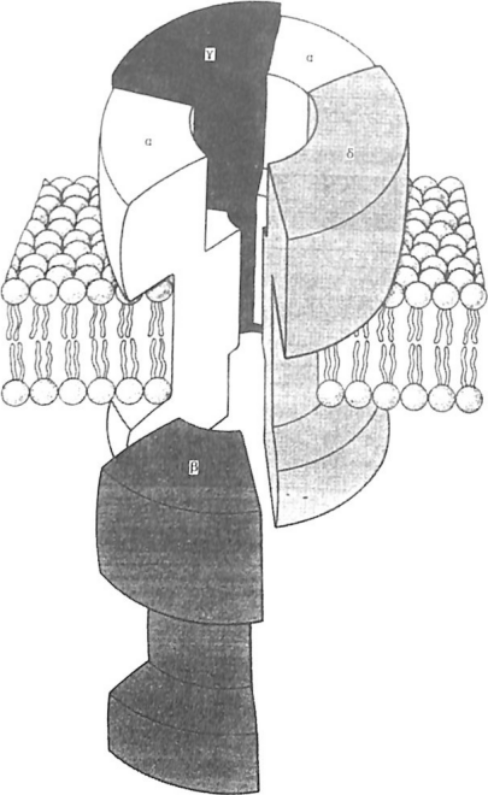
烟碱乙酰胆碱受体三维结构模型模式图
亚单位相结合而形成不活动的三倍体;当a亚单位与GTP相结合时,则该亚单位与b、r复合体相脱离,从而进一步调节各种突触后反应。
在某些情况下,由受体激活的G蛋白亚单位,可直接作用在例子通道而引起突触后电位,一些与乙酿胆碱结合的代谢趋向型受体。即属此类。这种受体起动突触后反应一般需数十毫秒、数秒甚至数分钟。而另G蛋白间接作用在离子通道上的受体则作用更慢。这种受体中,G蛋白要通过若干中间环节起作用,如腺苷酸环化酶和磷酸脂酶C;最近磷脂酶A2也被看作是效应器酶的一种。这些酶又开动细胞内的第二信使,引起进一步的生化反应。其中,腺苷酸环化酶专门结合在刺激型G蛋白上,它催化普遍存在的第二信使物质环鱗酸腺昔,cAMP可以直接作用于离子通道,但更多情况下是去激活蛋白激酶,如蛋白激酶A,它通过向蛋白质转移磷酸根而使离子通道的蛋白磷酸化从而引起通道的开放或关闭,产生突触后电位。磷脂酶C产生两种第二信使:肌醇三磷酸(IP3)和二酰基甘油(DAG)。IP3可以使细胞内储存的Ca2+释放而激活钾和氯离子通道,而DAG激活蛋白激酶C而调节不同离子通道。磷酸脂酶A2(phospholi pase A2) 则能引起细胞膜上的花生四烯酸的释放,从而进一步调节离子通道的活动。
上述这些受体引起的突触后反应一般在数毫秒到数分钟(有时为数小时)后出现,但作用最慢的和持续性长的递质有时并不激起突触后电位,它们通过改变基因的表达而间接地影响突触后细胞的电活动,使之反应加长。基因表达的改变可以由激活第二信使 并进而导致调节基因转录的蛋白磷酸化而达到,这种蛋白如cAMP反应元件结合蛋白 (CREB)的磷酸化就能调节某种特定基因的转录。例如,重复刺激某些胺能神经元可以增加去甲基肾上腺素合成的速率,这一效应受突触前能激活腺苷酸环化酶的代谢趋向型受体所调节。随后,蛋白激酶A活性增加则引起细胞核内转录调节因子(如CREB) 的磷酸化,从而决定其邻近的基因是否得到转录。在某些情况下,CREB的磷酸化可导致酪胺酸羟化酶合成增加,即引起儿茶酚胺类递质合成增多;在其他细胞内,这种转录调节因子可以引起合成离子通道或其他调节通道的蛋白质。
总而言之,神经递质改变细胞电活动的途径不仅是通过调节离子通道的状态,也通过调节通道及其相关蛋白质的合成。这种改变首先在几分钟到几小时内出现,但可持续数周、数月甚至更长,这是神经系统的长时程信息加工功能的物质基础。
关注心灵健康(预约) 0592—5515516
官方网站:心理健康 ◆ http://www.xmxljg.com
专家微博:心理资讯 ◆ http://weibo.com/fjxlzx/
企业微博:心灵鸡汤 ◆ http://e.weibo.com/xmxljg
心灵博客:心理辞典 ◆ http://blog.sina.com.cn/xmxlys
企业博客:心灵秘钥 ◆ http://blog.sina.com.cn/xmxljg
↓
厦门心理咨询(尊重您的个人私密,使隐私权得到充分的保障)
